presents
Coral Reef Bleaching
by Jason Buchheim
Director, Odyssey Expeditions
Copyright 1998-2013, Odyssey Expeditions- Marine Biology Learning Center Publications
Permission granted to cite paper, but not copy entire document!
presents
Coral Reef Bleaching
by Jason Buchheim
Director, Odyssey Expeditions
Copyright 1998-2013, Odyssey Expeditions- Marine Biology Learning Center Publications
Permission granted to cite paper, but not copy entire document!
Coral reef bleaching, the whitening of diverse invertebrate taxa, results from the loss of symbiotic zooxantheallae and/or a reduction in photosynthetic pigment concentrations in zooxanthellae residing within scleractinian corals. Coral reef bleaching is caused by various anthropogenic and natural variations in the reef environment including sea temperature, solar irradiance, sedimentation, xenobiotics, subaerial exposure, inorganic nutrients, freshwater dilution, and epizootics. Coral bleaching events have been increasing in both frequency and extent worldwide in the past 20 years. Global climate change may play a role in the increase in coral bleaching events, and could cause the destruction of major reef tracts and the extinction of many coral species.
| Introduction
Coral reefs are found in circumtropical shallow tropical waters along the shores of islands and continents. The reef substrate is mainly composed of calcium carbonate from living and dead scleractinian corals. Many other invertebrates, vertebrates, and plants live in close association to the scleractinian corals, with tight resource coupling and recycling, allowing coral reefs to have extremely high productivity and biodiversity, such that they are referred to as the Tropical Rainforests of the Oceans. Corals live in very nutrient poor waters and have certain zones of tolerance to water temperature, salinity, UV radiation, opacity, and nutrient quantities. Scleractinian corals build skeletons of calcium carbonate sequestered from the water. When the coral polyp dies, this skeleton remains incorporated in the reef framework. Scleractinian corals are in the Phylum Cnidaria, and they receive their nutrient and energy resources in two ways. They use the traditional cnidarian strategy of capturing tiny planktonic organisms with their nematocyst capped tentacles, as well as having a obligate symbiotic relationship with a single cell algae known as zooxanthellae. Zooxanthellae are autorophic microalgaes belonging to various taxa in the Phylum Dinoflagellata. Zooxanthellae live symbiotically within the coral polyp tissues and assist the coral in nutrient production through its photosynthetic activities. These activities provide the coral with fixed carbon compounds for energy, enhance calcification ,and mediate elemental nutrient flux. The host coral polyp in return provides its zooxanthellae with a protected environment to live within, and a steady supply of carbon dioxide for its photosynthetic processes. The symbiotic relationship allows the slow growing corals to compete with the faster growing multicellular algaes because the tight coupling of resources and the fact that the corals can feed by day through photosynthesis and by night through predation. The tissues of corals themselves are actually not the beautiful colors of the coral reef, but are instead clear. The corals receive their coloration from the zooxanthellae living within their tissues. Coral reef ecosystems world-wide have been subject to unprecedented degradation over the past few decades. Disturbances affecting coral reefs include anthropogenic and natural events. Recent accelerated coral reef decline seems to be related mostly to anthropogenic impacts (overexploitation, overfishing, increased sedimentation and nutrient overloading. Natural disturbances which cause damage to coral reefs include violent storms, flooding, high and low temperature extremes, El Nino Southern Oscillation (ENSO) events, subaerial exposures, predatory outbreaks and epizootics. Coral reef bleaching is a common stress response of corals to many of the various disturbances mentioned above. Beginning in the 1980s, the frequency and widespread distribution of reported coral reef bleaching events increased. Widespread bleaching, involving major coral reef regions and resulting in mass coral mortality has raised concerns about linkage of the events to global phenomenons including global warming or climate change and increased UV radiation from ozone depletion. This paper examines the causes of coral reef bleaching and addresses the impact of global climate change on coral reefs. Coral reef bleaching Bleaching, or the paling of zooxanthellate invertebrates,
occurs when (i) the densities of zooxanthellae decline and / or (ii) the
concentration of photosynthetic pigments within the zooxanthellae fall
(Kleppel et al. 1989). Most reef-building corals normally contain
around 1-5 x 106 zooxanthellae cm-2 of live surface
tissue and 2-10 pg of chlorophyll a per zooxanthella. When corals bleach
they commonly lose 60-90% of their zooxanthellae and each zooxanthella
may lose 50-80% of its photosynthetic pigments (Glynn 1996). The pale appearance
of bleached scleractinian corals and hydrocorals is due to the cnidarians
calcareous skeleton showing through the translucent tissues (that are nearly
devoid of pigmented zooxanthellae).
Three hypotheses have been advanced to explain the cellular mechanism of bleaching, and all are based on extreme sea temperatures as one of the causative factors. High temperature and irradiance stressors have been implicated in the disruption of enzyme systems in zooxanthellae that offer protection against oxygen toxicity. Photosynthesis pathways in zooxanthallae are impaired at temperatures above 30 degrees C, this effect could activate the disassociation of coral / algal symbiosis. Low- or high-temperature shocks results in zooxanthellae low as a result of cell adhesion dysfunction. This involves the detachment of cnidarian endodermal cells with their zooxanthellae and the eventual expulsion of both cell types. It has been hypothesized that bleaching is an adaptive mechanism which allows the coral to be repopulated with a different type of zooxanthellae, possibly conferring greater stress resistance. Different strains of zooxanthellae exist both between and within different species of coral hosts, and the different strains of algae show varied physiological responses to both temperature and irradiance exposure. The coral / algal association may have the scope to adapt within a corals lifetime. Such adaptations could be either genetic or phenotypic.
Ecological causes of coral bleaching As coral reef bleaching is a general response to stress, it can be induced by a variety of factors, alone or in combination. It is therefore difficult to unequivocally identify the causes for bleaching events. The following stressors have been implicated in coral reef bleaching events. Temperature Coral species live within a relatively narrow temperature
margin, and anomalously low and high sea temperatures can induce coral
bleaching. Bleaching events occur during sudden temperature drops accompanying
intense upwelling episodes, (-3 degrees C to 5 degrees C for 5-10 days),
seasonal cold-air outbreaks. Bleaching is much more frequently reported
from elevated se water temperature. A small positive anomaly of 1-2 degrees
C for 5-10 weeks during the summer season will usually induce bleaching.
Bleaching during the summer months, during seasonal temperature and irradiance maxima often occurs disproportionately in shallow-living corals and on the exposed summits of colonies. Solar radiation has been suspected to play a role in coral bleaching. Both photosyntheticaly active radiation (PAR, 400-700nm) and ultraviolet radiation (UVR, 280-400nm) have been implicated in bleaching. Subaerial Exposure Sudden exposure of reef flat corals to the atmosphere during events such as extreme low tides, ENSO-related sea level drops or tectonic uplift can potentially induce bleaching. The consequent exposure to high or low temperatures, increased solar radiation, desiccation, and sea water dilution by heavy rains could all play a role in zooxanthellae loss, but could also very well lead to coral death. Sedimentation Relatively few instances of coral bleaching have been linked solely to sediment. It is possible, but has not been demonstrated, that sediment loading could make zooxanthellate species more likely to bleach. Fresh Water Dilution Rapid dilution of reef waters from storm-generated precipitation and runoff has been demonstrated to cause coral reef bleaching. Generally, such bleaching events are rare and confined to relatively small, nearshore areas. Inorganic Nutrients Rather than causing coral reef bleaching, an increase in ambient elemental nutrient concentrations (e.g. ammonia and nitrate) actually increases zooxanthellae densities 2-3 times. Although eutrophication is not directly involved in zooxanthellae loss, it could cause secondary adverse affects such as lowering of coral resistance and greater susceptibility to diseases. Xenobiotics Zooxanthellae loss occurs during exposure of coral to elevated concentrations of various chemical contaminants, such as Cu, herbicides and oil. Because high concentrations of xenobiotics are required to induce zooxanthellae loss, bleaching from such sources is usually extremely localized and / or transitory . Epizootics Pathogen induced bleaching is different from other sorts of bleaching. Most coral diseases cause patchy or whole colony death and sloughing of soft tissues, resulting in a white skeleton (not to be confused with bleached corals). A few pathogens have been identified the cause translucent white tissues, a protozoan.
Spatial and temporal range of coral reef bleaching Mass coral moralities in coral reef ecosystems have been reported in all major reef provinces since the 1870s. The frequency and scale of bleaching disturbances has increased dramatically since the late 70s. This is possibly due to more observers and a greater interest in reporting in recent years. More than 60 coral reef bleaching events out of 105 mass coral moralities were reported between 1979-1990, compared with only three bleaching events among 63 mass coral moralities recorded during the preceding 103 years. Nearly all of the worlds major coral reef regions (Caribbean/
western Atlantic, eastern Pacific, central and western Pacific, Indian
Ocean, Arabian Gulf, Red Sea) experienced some degree of coral bleaching
and mortality during the 1980s.
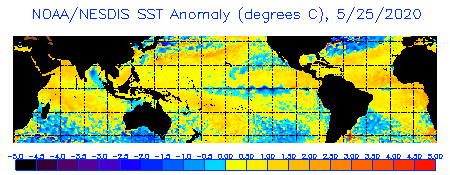
Prior to the 1980s, most mass coral moralities were related to non-thermal disturbances such as storms, aerial exposures during extreme low tides, and Acanthaster outbreaks. Coral bleaching accompanied some of the mortality events prior to the 1980s during periods of elevated sea water temperature, but these disturbances were geographically isolated and restricted to particular reefs zones. In contrast, many of the coral bleaching events observed in the 1980s occurred over large geographic regions and at all depths. Most of the coral reef bleaching events of the 1980s occurred during years of large-scale ENSO activity. Global change and reef bleaching Of the causing stressors of coral reef bleaching, many are related to local environmental degradation and reef overexploitation. Of the stressors mentioned above, only sea water temperature and solar irradiance have possible global factors driving changes and extremes. Global warming, along with ENSO events, change sea water temperatures. Ozone depletion increases the amount of UVR reaching the Earths surface, and possibly causing coral bleaching events. Increased sea temperatures and solar radiation (especially UV radiation), either separately or in combination, have received consideration as plausible large-scale stressors. In most instances, wherever coral reef bleaching was reported, it occurred during the summer season or near the end of a protracted warming period. Coral bleaching was reported to have occurred during periods of low wind velocity, clear skies, calm seas and low turbidity, when conditions favor localized heating and high penetration of short wave length (UV) radiation. Also less oxygen is held by water at higher temperatures. Potentially stressful high sea temperatures and UV radiation flux could conceivably cause coral reef bleaching on a global scale with suspected greenhouse warming and the thinning of the ozone layer. As reef building corals live near their upper thermal
tolerance limits, small increases in sea temperature (.5 1.5 degrees C)
over several weeks or large increases (3-4 degrees C) over a few days will
lead to coral dysfunction and death. Anomalously high sea temperatures
have often been reported in the Caribbean-wide series of bleaching events
that occurred during 1986-88, leading to hypothesis that global warming
was having an effect on the coral reefs in this region.
Solar ultraviolet radiation is potentially harmful to reef corals and their symbiotic. UV radiation can readily penetrate clear sea water, and reef building corals contain UV-absorbing compounds capable of blocking potentially damaging UV radiation. These compounds are produced in response to ambient UV levels and the concentration in corals is usually an inverse function of depth, but it is not known if bleaching responses are related to variations in UV flux that exceed the protective capacity of UV-absorbing compounds. There is a possible interaction between temperature and UV, with temperature significantly reducing zooxanthelae densities and also the concentration of UV absorbing compounds in a reef zooanthid, thus potentially increasing the exposure of the symbionts to the direct effects of UV radiation.
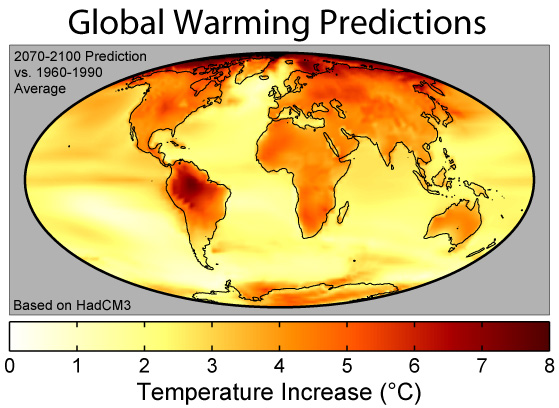
Conclusion If a global warming trend impacts on shallow tropical and subtropical seas, we may expect an increase in the frequency, severity and scale of coral reef bleaching. Coral mortality could exceed 95% regionally with species extirpation and extinctions. A conservative temperature increase of 1-2 degrees C would cause regions between 20-30 degrees N to experience sustained warming that falls within the lethal limits of most reef-building coral species. In conjunction with sea temperature rise would be a sea level rise, and it has been suggested that sea level rise would suppress coral growth or kill many corals through drowning or lower light levels. Some coral populations and their endosymbiotic zooxanthellae may be able to adapt to the extreme conditions predicted during global climate change. Refuges in benign habitats, such as deep, sunlit reef substrates, oceanic shoals and relatively high latitude locations, might exist, but widespread coral mortality and reef decline would be expected in shallow reef zones in most low latitude. Even if significant sea warming and elevated irradiance levels do not occur, coral reef degradation from anthropogenic pollution and overexploitation will still continue, a result of unrelenting human population growth. |
|
References Atwood DK, Sylvester JC, Corredor JE, Morell JM, Mendez A, Nodal WJ, Huss BE, Foltz C (1988) Sea surface temperature anomalies for the Caribbean, Gulf of Mexico, Florida reef tract and the Bahamas considered in light of the 1987 regional coral bleaching event. Proc. Assoc. Mar. Lab. Caribbean 27:47. Barnes DJ, Chalker BE (1990) Calcification and photosynthesis in reef-building corals and algae. In Coral Reefs (ed Dubinsky Z), pp. 109-131. Ecosystems of the World 25, Elsevier, Amsterdam. Blumthaler M, Amback W (1990) Indication of increasing solar UV-B radiation and flux in Alpine regions. Science 248:206-208. Bradshaw AD, Hardwick K (1989) Evolution and stress genotypic and phenotypic components. J. Linn. Soc 37:137-155. Brown BE (1987) Worldwide death of corals-natural cyclical events or man-made pollution? Marine Pollution Bulletin 18:12-13. Brown BE, Howard LS (1985) Assessing the effects of stress on reef corals. Advances in Marine Biology, 22:1-63. Brown BE, Ogden JC (1993) Coral bleaching. Scientific American 268:64-70. Buddemier RW, Fuatin DG (1993) Coral bleaching relative to elevated seawater temperature in the Andaman Sea (Indian Ocean) over the last 50 years. Coral Reefs 15: 151-152. Coffroth MA, Lasker HR, Oliver JK (1990) Coral mortality outside of the eastern Pacific during 1982-1983; relationship to El Nino. In: Glynn PW9ed) Global ecological consequences of the 1982-83 El Nino-Southern Oscillation. Elsevier, Amsterdam, pp. 141-182. Coles SL, Jokeil PL, Lewis CR (1976) thermal tolerance in tropical versus subtropical Pacific reef corals. Pac. Sci. 30:159-166. Coles SL, Fadlallah YH (1991) Reef coral survival and mortality at low temperatures in the Arabian Gulf: new species-specific lower temperature limits. Coral Reefs 9:231-237. DElia CF, Wiebe WJ (1990) Biogeochemical nutrient cycles in coral-reef ecosystems. In: Coral Reefs (ed Dubinsky Z), pp. 49-74. Ecosystems of the World 25, Elsevier, Amsterdam. DElia CF, Buddemeier RW, Smith SV (1991) Workshop on coral bleaching, coral reef ecosystems and global change; report of proceedings. Maryland Sea Grant Coll. Publ. No. UM-SG-TS-91-03, College Park, University of Maryland, Md. Dubinsky Z, Stambler N (1996) Eutrophication, marine pollution and coral reefs. Global Change Biology 2:511-526. Dunlap WC, Chalker BE (1986) Identification and quantification of near-UV absorbing compounds (S-320) in a hermatypic scleractinian. Coral Reefs 5:155-159. Dunlap WC, Chalker BE, Oliver JK (1986) Bathymetric adaptations of reef-building corals at Davies Reef, Great Barrier Reef, Australia. III. UV- B absorbing compounds. J Exp Mar Biol Ecol, 104:239-248. Egana AC, DiSalvo LH (1982) Mass expulsion of zooxanthellae by Easter Island corals. Pacific Science 36:61-63. Fleischmann EM (1989) The measurement and penetration of ultraviolet radiation into tropical marine water. Limnol. Oceanogr. 34:1623-1629. Ginsburn RH (compiler) (1994) Proceedings of the Colloguium on Global Aspects of Coral Reefs: Health, Hazards and History, 1993. Rosentiel School of Marine and Atmospheric Science, University of Miami, 420 pp. Gleason DF, Wellignton GM (1993) Ultraviolet radiation and coral bleaching. Nature 365:836-838. Gleason DF, Wellington GM (1995) variation in UVB sensitivity of planula larvae of the coral Agaricia agaricites along a depth gradient. Marine Biology 123:693-703. Glynn PW (1984) Widespread coral mortality and the 1982/83 El Nino warming event. Environmental Conservation 11:133-146. Glynn PW (1988) Coral bleaching and mortality in the tropical eastern Pacific during the 1982-83 El Nino warming event. In: Ogden J, Wicklund R (eds) Mass bleaching of coral reefs in the Caribbean: a research strategy. NOAAs Undersea Res Prog, St Croix, US Virgin Islands, Res Rpt 88-2:42-45. Glynn PW (1991) Coral reef bleaching in the4 1980s and possible connections with global warming. Trends in Ecology and Evolution 6:175-179. Glynn PW (1993) Coral reef bleaching: ecological perspectives. Coral Reefs 12:1-17. Glynn PW (1996) Coral reef bleaching: facts, hypotheses and implications. Global Change Biology 2:495-509. Glynn PW, DCrox L (1990) Experimental evidence for high temperature stress as the cause of El Nino-coincident coral mortality. Coral Reefs 8:181-191. Glynn PW, Howard LS, Corcoran E, Freay AD (1984) The occurrence and toxicity of herbicides in reef-building corals. Marine Pollution Bulletin 15:370-374. Goreau TF (1964) Mass expulsion of zooxanthellae from Jamaican reef communities after Hurricane Flora. Science 145:383-386. Goreau TJ, Hayes RL (1994) Coral bleaching and ocean hot spots. Ambio 23:176-180. Grigg RW, Dollar SJ (1990) Natural and anthropogenic disturbance on coral reefs. In: Coral Reefs (ed. Dubinsky Z), pp. 439-452. Elsevier, Amsterdam. Hughes T (ed.) (1993) Disturbance: effects on coral reef dynamics. Coral Reefs 12:115-234. Iglesias-Prieto R, Matta JL, Robins WA, trench RK (1992) Photsynthetic responses to elevated temperature in the symbiotic dinoflagellate Symbiodinium microadriaticum in culture. Proceedings of the National Academy of Science USA 89:10302-10305. Jackson JBC, Cubit JD, Keller BD (1989) Ecological effects of a major oil spill on Panamanian coastal marine communities. Science 243:37-44. Jaap WC (1988) The 1987 zooxanthellae expulsion event at Florida reefs. NOAAs Undersea Res Prog Res Rpt 88:24-29. Jokiel PL, Coles SL (1990) Response of Hawaiian and other Indo-Pacific reef corals to elevated temperature. Coral Reefs 8:155-162. Kleppel GS, Dodge RE, Reese CJ (1989) Changes in pigmentation associated with the bleaching of stony corals. Limnology and Oceanography 34:1331-1335. Kushmaro A, Loya Y, Fine M, Rosenberg E (1996) Bacterial infection and coral bleaching. Nature 380:396. Lesser MP, Shick JM (1989) Effects of irradiance and ultraviolet radiation on photoadaptation in zooxanthellae of Aiptasia pallida: primary production, photoinhibition, and enzymic defenses against oxygen toxicity. Mar Biol 102:243-255. Lesser MP, Shick JM (1990) effects of visible and ultraviolet radiation on the ultrastructure of zooxanthellae (Symbiodinium sp.) in culture and in situ. Cell Tissue Res 261:501-508. Lesser MP, Stochaj WR, Tapley DW, Shick JM (1990) Bleaching in coral reef anthozoans: effects of irradiance, ultraviolet radiation, and temperature on the activities of protective enzymes against active oxygen. Coral Reefs 8:225-232. Muscatine L (1990) the role of symbiotic algae in carbon and energy flux in reef corals. In: Coral Reefs (ed Dubinsky Z), pp. 75-87. Ecosystems of the World 25, Elsevier, Amsterdam. Pearson RG (1981) Recovery and recolonization of coral reefs. Marine Ecology Progress Series 4:105-122. Stimson J, Kinzie III RA (1991) The temporal pattern and rate of release of zooxanthellae from the reef coral Pocillopora damicornis (Linnaeus) under nitrogen-enrichment and control conditions. J Exp Mar Bio Ecol 153:63-74. Tsonis AA, Elsner JM (1989) Testing the global warming hypothesis, Geophys Res Lett 16:795-797. Upton SJ, Peters EC (1986) A new and unusual species of coccidium (Apicomlexa: Agammococcidorida) from Caribbean scleractinian corals. Journal of Invertebrate Pathology 47:184-193. Ware JR, Fuatin DG, Buddemeir RW (1996) Patterns of coral bleaching: modeling the adaptive bleaching hypothesis. Ecol. Model 84:199-214. Williams EH, Bunkley-Williams L (1990) The world-wide coral bleaching cycle and related sources of coral mortality. Atoll Research Bulletin 335:1-71. |
Marine Biology resources by Odyssey Expeditions Tropical Marine Biology Voyages | ||
|